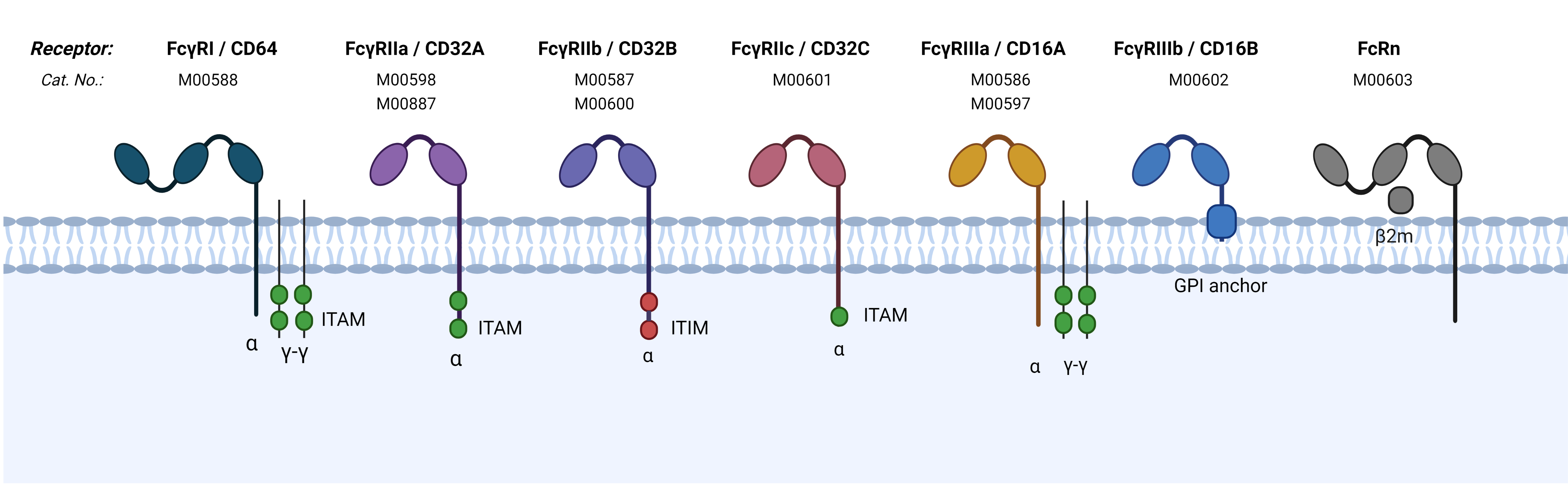



































GenScriptは、全てのIgGサブクラスに対する結合評価をカバーできるFcγRおよびFcRn過剰発現細胞株のラインナップを提供しています。これらの細胞株は、Fc-FcγRおよびFc-FcRn結合親和性評価のためのセルベース結合アッセイ用にデザインされています。
病原体あるいは感染細胞に結合した抗体のFc領域と結合した後、FcγRsは抗体依存性細胞貪食(ADCP)または抗体依存性細胞傷害(ADCC)を含むエフェクター機能を介在します。抗体をベースとした医薬品開発に関するFcエンジニアリングにおいて、Fc-FcγR結合親和性の上昇または低下が、エフェクター機能を強化または弱化する可能性があります。FcRnは血清中のIgG型抗体のリサイクルに関わっており、抗体の安定性と半減期を高めます。そのため、Fc-FcRn結合親和性を増強するための抗体Fcエンジニアリングは、抗体の循環半減期を伸ばす可能性があります。
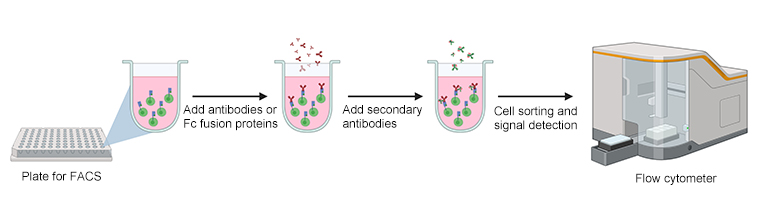
図1:フローサイトメーターを使用した、Fc受容体発現細胞ベースの結合アッセイ
フローサイトメーターを使用したセルベース結合アッセイは、結合親和性を評価する方法の1つです。
一塩基多型は、抗体ベースの薬剤または免疫療法に対する個人の免疫反応に影響する、FcγR多型に関与します。同一の抗体ベースの薬剤でも、FcγR多型を持つ患者では異なる効果となる可能性があります。GenScriptは、多型研究を行うためのFcγR多型過剰発現細胞株を提供しています。
表1:ヒトFcRγおよびFcRn 受容体の、免疫グロブリンサブクラスに対する結合性および機能
| 名称 | 別名 | 多型 | IgGサブクラスに対する結合性 | 機能 |
|---|---|---|---|---|
| FcγRIIIa | CD16A | V158 | Higher affinity to all human IgGs than F158 | Activation /Inhibition |
| F158 | Lower affinity to all human IgGs than V158 | Activation /Inhibition | ||
| FcγRIIIb | CD16B | - | IgG1,IgG3 | Activation |
| FcγRIIa | CD32A | H131 | IgG1,IgG2,IgG3,IgG4 | Activation /Inhibition |
| R131 | lower affinity to IgG1 and IgG2 than H131, IgG3, IgG4 | Activation /Inhibition | ||
| FcγRIIb | CD32B | I232 | IgG1,IgG2,IgG3,IgG4 | Inhibition |
| T232 | IgG1,IgG2,IgG3,IgG4 | Inhibition | ||
| FcγRIIc | CD32C | - | IgG1,IgG2,IgG3,IgG4 | Activation |
| FcγRI | CD64 | - | IgG1,IgG3,IgG4 | Activation |
| FcRn | FcRn | - | IgG1,IgG2,IgG3,IgG4 | Recycling, transport, uptake |